Background / Past activities
How we started:
Immune competence of murine glial cells (1995-2001)
Following injury in the CNS, astrocytes and microglia show an immediate response in the form of proliferation, cytokine production and expression of immunocompetent molecules. Our objectives were to better understand the role of these glial cells in the context of inflammation and/or immune response in the central nervous system. In an effort to analyze the mechanisms that underlie their immune function, following studies were undertaken. Characterization of the capacity of primary murine astrocytes and microglia for antigen uptake and processing led us to the discovery that astrocytes as well as microglia express the mannose receptor, a pattern-recognition receptor involved in innate and adaptive immunity (1-4). Comparison of the molecular machinery that underlies MHC class II trafficking in immunocompetent astrocytes and microglia indicated major differences in the capacity of these glial cells to process the invariant chain and to express active cathepsin L and cathepsin S that are mandatory for antigen processing (5). Using defined combinations of cytokines to simulate in vitro different immune environments observed in disease or inflammation, we observed that astrocytes down-regulate their T cell stimulation and MHC II trafficking functions while microglia maintain their antigen presentation potential (6). Whereas the classical MHC I and MCH II molecules bind to protein antigens, the non-classical MHC CD1 molecules bind to lipid and glycolipid antigens and as such may present self glycolipids derived from brain tissue during the local inflammatory reaction and demyelination. Ex-vivo analysis of CD1 expression by resident and infiltrating cells isolated from brains of healthy mice and mice undergoing experimental autoimmune encephalomyelitis indicated a differential regulation of expression by microglia, macrophages and T cells in the central nervous system upon inflammation and demyelination (7).
In summary, these studies contributed a set of novel data on the relevance of the cytokine environment for the control of molecules involved in antigen uptake (mannose receptor) and MHC II expression and trafficking (cathepsins) that were found to differ between astrocytes and microglia.
1-BURUDI, E.M.E. et al. (1999) Identification and functional characterization of the mannose receptor in astrocytes. Glia, 25, 44-55 PMID9888297
2-BURUDI, E.M.E., and REGNIER-VIGOUROUX, A. (2001) Regional and cellular expression of mannose receptor in the post-natal developing mouse brain. Cell Tissue Res, 303, 307-317 PMID11320646
3-ZIMMER, H. et al. (2003) Functional characterization of mannose receptor expressed by immunocompetent mouse microglia. Glia, 42, 89-100 PMID12594740
4-REGNIER-VIGOUROUX, A. (2003) The mannose receptor in the brain. International Review of Cytology, 226, 321-342 PMID12921240
5-GRESSER, O. et al. (2001) Immunocompetent astrocytes and microglia display major differences in the processing of the invariant chain and in the expression of active Cathepsin L and Cathepsin S. Eur. J. Immunol., 31, 1813-1824 PMID11433378
6-GRESSER, O. et al. (2000) Tumor necrosis alpha and interleukin-1 alpha inhibit through different pathways interferon-gamma-induced antigen presentation, processing and MHC class II surface expression on astrocytes, but not on microglia. Cell Tissue Res, 300, 373-382 PMID10928267
7-BUSSHOFF, U. et al. (2001) CD1 expression is differentially regulated by microglia, macrophages and T cells in the CNS upon inflammation and demyelination. J. Neuroimmunol., 113, 220-230 PMID11164905
How it developed: Microglia and brain cancer (2001-2015)
Microglia-mediated anti-tumour activities and parvovirus
Specificity and safety of use of the parvovirus as a vector for brain gene therapy
The safe use of parvovirus for glioma therapy was analysed in a proof-of-the principle study in the mouse. Our data (Abschuetz et al, 2006; Nickles and Régnier-Vigouroux, unpublished data) indicate that, in the natural host, a majority of normal glial cells (astrocytes and microglia) are not competent for parvovirus replication and that the abortive infection taking place in a minor fraction of these cells fails to impede their survival and immunocompetence. Similar experiments conducted with human microglia demonstrated a lack of parvovirus toxicity towards these cells (Kees and Régnier-Vigouroux, unpublished data). These features argue for the safety of parvovirus-based therapies of brain tumours that may harbour reactive and proliferative glial cells. For more information about parvovirus-based therapies of glioblastoma, we refer the reader to the published work of our colleagues, Christiane Dinsart, Jürg Nuesch and Jean Rommelaere (Tumor Virology Division, DKFZ, Heidelberg, Germany).
Characterisation of microglia pro- and anti-tumour activities
1-Tumor-supporting microglia can be turned into efficient oncotoxic cells
Whereas the pro-tumour activities of murine microglia are well documented, those of human cells were still scarce when we started this project. We modified available protocols to obtain human microglia, isolated from brain tumour specimen, and analysed them for their pro- and anti-tumor activities (collaboration with Dr. Christel Herold-Mende, Neurosurgery, Heidelberg). We observed that untreated, i.e. “resting” microglia secrete factors that support tumour growth and migration in a 3D collagen invasion assay. Stimulation of human microglia with a TLR-3 agonist however led to an anti-tumour phenotype characterised by reduction of tumour cell growth and migration and by cytotoxicity. Microglia toxicity was specifically directed at tumor cells, as human astrocytes and neurons were not affected by the supernatant of stimulated microglia. These data indicate that human microglia isolated from tumour tissue have a tumour-supporting phenotype that can be switched to an anti-tumour phenotype by application of (a) proper stimulus/stimuli. However, these anti-tumour activities were not induced when microglia activation was performed in the presence of human glioma cells, indicating a strong control exerted by glioma cells on the regulation of microglia activation state (Kees et al, 2012).
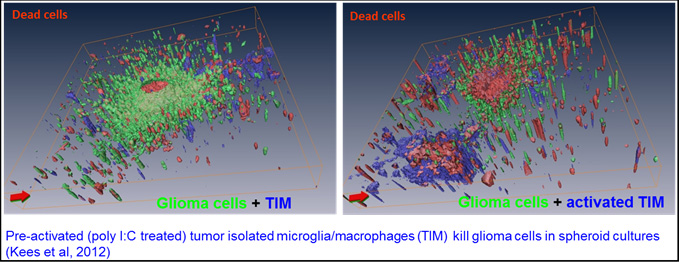
2-Characterisation of microglia oncotoxicity
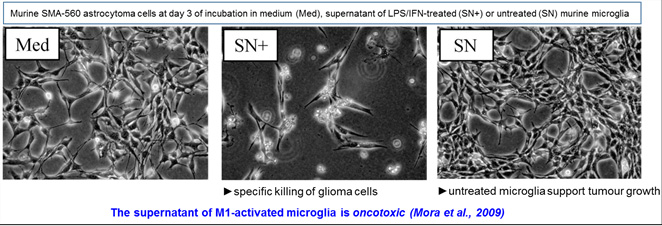
Very little progress has been made in understanding the molecular basis of microglia cytotoxicity since its first characterisation towards a non-brain tumour cell line. We reported the first detailed characterisation of the lipopolysaccharide plus interferon-gamma-induced cytotoxic activity of primary microglia towards glioma cell lines in the mouse system (Mora et al, 2009). Stimulated microglia secreted proteic factors that efficiently killed TNF-a and TRAIL-resistance glioma cell lines without affecting the survival of primary cultures of astrocytes or neurons. Cell death was autophagy-dependent and resulted from a blockade of the basal autophagic flux present in tumour cells. These observations demonstrate that glioma cells resistant to apoptotic death ligands could be successfully and specifically killed through autophagy-dependent death induced by appropriately activated microglia. We could mimic part of these effects through the chemical inhibition of sphingosine kinase in murine and human glioma cells, supporting the promising use of sphingosine kinase inhibitors in glioma therapy (Mora et al., 2010).
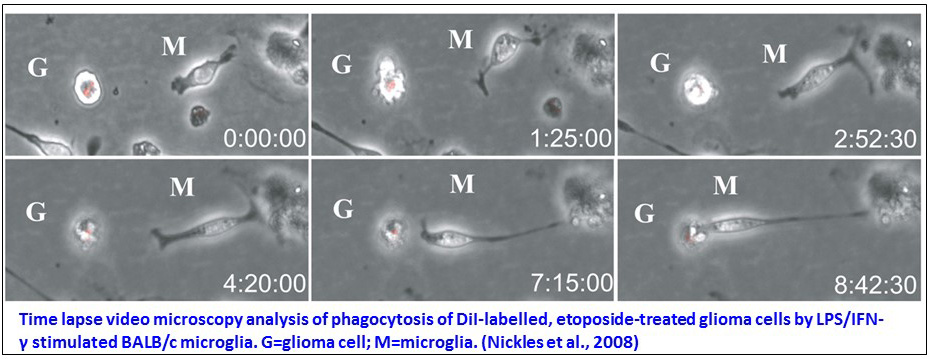
We furthermore analysed the microglia phagocytic activity for the removal of apoptotic glioma cells that had hardly been analysed in spite of its relevance to tissue damage prevention. Our comparative analysis of microglia originating from three different mice strains evidenced a phosphatidylserine-independent clearance of murine glioma cells at an advanced stage of death, suggesting microglia recognition of late apoptotic markers. This phagocytic activity was restricted to a microglia subset representing 30 to 70% of the population according to the used strain. Expression of receptors involved in late apoptotic markers recognition therefore seems confined to a subpopulation of microglia and to be strain-dependent (Nickles et al., 2008).
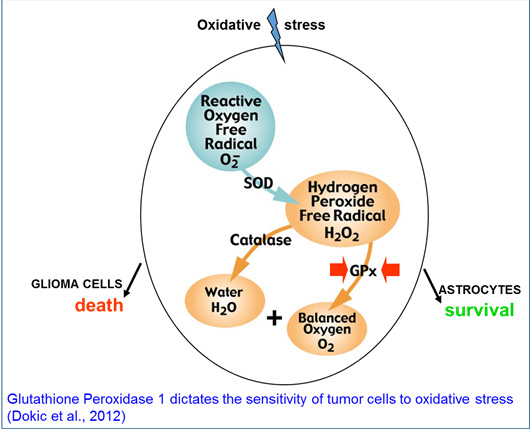
3-Glioblastoma resistance to therapy:
In the course of the above mentioned studies, we observed that normal astrocytes and glioblastoma cells kept in culture differed in their response to oxidative stress, suggesting differences in the detoxification potential of these cells. We indeed identified the glutathione peroxidase 1 enzyme to be a crucial element over other antioxidant enzymes for oxidative stress regulation in murine and human glioblastoma cells. Our data suggest that mapping the antioxidant enzyme status of glioblastoma may prove to be a useful tool for personalized ROS/RNS inducing therapies (Dokic et al, 2012).
This study prompted us, in collaboration with Klaus-Josef Weber (Heidelberg University Hospital, Department of Radiation Oncology and Radiation), to further analyse two patient-derived glioblastoma cell lines in order to investigate chromosomal instability and radiation response mechanisms in glioblastoma cells. The cell resistance to low and high linear energy transfer (LET) radiation was assessed using clonogenic survival assay and their intrinsic chromosome instability status using fluorescence in situ hybridization. DNA damage was analyzed by pulsed-field gel electrophoresis and by γ-H2AX foci quantification. Expression of DNA damage response proteins was assessed by immunoblot. We observed an increased radioresistance to X-rays as well as carbon ions in glioblastoma cells exhibiting high levels of naturally occuring chromosomal instability and impaired ATM signaling, as reflected by lack of phosphorylation of ATM, CHK2 and p53 after double-strand breaks induction. These results indicate the existence of highly radioresistant glioblastoma cells, characterized by dysfunctional ATM signaling and high levels of intrinsic chromosomal instability (Dokic et al, 2014).
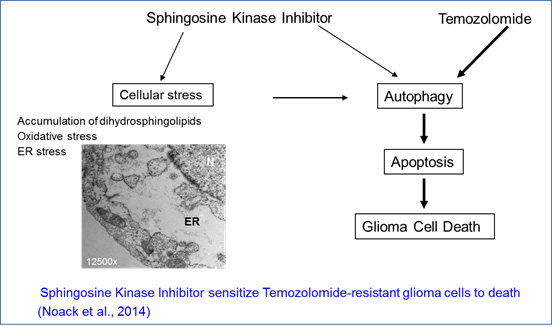
Given the specific killing of glioma cells by the sphingosine kinase inhibitor, we investigated the effects of combining sphingosine kinase inhibitors with temozolomide to improve the efficacy and efficiency of these drugs towards tumor cells. We observed that combination of sub-lethal doses of each drug very efficienly and specifically killed temozolomide-resistant human glioblastoma cell lines. The drug combination induced an endoplasmic reticulum stress, consecutive to disruption of sphingolipid metabolism, that led to induction of autophagy and thereafter apoptosis (Noack et al, 2014).
References
ABSCHUETZ, A. et al. (2006) The oncolytic murine autonomous parvovirus, a candidate vector for glioma gene therapy, is innocuous to normal and immunocompetent mouse glial cells. Cell Tissue Res, 325, 423-436
NICKLES, D., et al. (2008) End-stage dying glioma cells are engulfed by mouse microglia with a strain-dependent efficacy. J. Neuroimmunol., 197,10–20
MORA, R., et al. (2009) TNF-α- and TRAIL-resistant glioma cells undergo autophagy-dependent cell death induced by activated microglia. Glia, 57, 561-581.
MORA, R., et al. (2010) Sphingolipid rheostat alterations related to transformation can be exploited for specific induction of lysosomal cell death in murine and human glioma. Glia, 58, 1364-1383
KEES, T., et al. (2012) Microglia isolated from glioma patients gain anti-tumor activities upon poly (I:C) stimulation. Neuro-Oncology, 14, 64-78. Epub 2011 Oct 20
DOKIC, I., et al. (2012) Glutathione peroxidase 1 activity dictates the sensitivity of glioblastoma cells to oxidative stress. Glia, 60, 1785-1800. Epub 2012 Aug 1
DOKIC, I., et al. (2014) High resistance to X-rays and therapeutic carbon ions in glioblastoma cells bearing dysfunctional ATM associates with intrinsic chromosomal instability Int J Radiat Biol, Sep 8:1-9 [Epub ahead of print]
NOACK, J., et al. (2014) A sphingosine kinase inhibitor combined with temozolomide induces glioblastoma cell death through accumulation of dihydrosphingosine and dihydroceramide, endoplasmic reticulum stress and autophagy. Cell Death and Disease, 5, e1425; doi:10.1038/cddis.2014.384